【堀先生の部屋】シラカンバについて
樹木生態研究会最高顧問 堀 大才

天然分布
シラカンバ(白樺)は樹高20m以上になるカバノキ科カバノキ属Betulaの喬木性落葉広葉樹である。世界にはカバノキ属は40種ほど、変種も入れると50種ほどが数えられており、日本には10から12種ほどが天然に自生している(研究者によって若干種数が異なる)。シラカンバB. platyphylla var. japonica と基本種のコウアンシラカンバB. platyphylla var. platyphylla、近縁種のオウシュウシラカンバB. pendulaおよびアメリカシラカンバB. papyriferaなどをひとくくりにして広義のシラカンバ類とみなすと、広義のシラカンバ類は北半球の寒冷な地方では最も個体数の多い周北極要素の落葉広葉樹である。日本産シラカンバの基本種であるコウアンシラカンバ(興安白樺)はバイカル湖以東のシベリア東部や中国東北地方の森林地帯で最も普通に見られ、オウシュウシラカンバ(欧州白樺)はシベリア全域からヨーロッパにまで分布する。アメリカシラカンバは北米大陸北部に分布する。シラカンバ類は冷温帯や亜寒帯、あるいは山地帯や亜高山帯の森林の林縁や崩壊地でごく普通に見られ、厳冬期の耐凍性は−35℃に達するとされるほど耐寒性が極めて強いので、森林限界を越えた寒帯や高山帯にも生育することがある。
日本産シラカンバの天然分布は、北は北海道全域から、本州の日本海側では福井県の山地帯まで、太平洋側では静岡県の山地帯までである。福井県と静岡県の間に位置する岐阜県の山岳地帯にも分布する。
北海道では標高の低い平地にも生育するが、南に下るにつれて分布の標高が高くなり、本州の関東地方や中部地方では低いところでも約600m以上、高いところでは1600m以上となる。
近畿地方から九州地方には存在しないが、近年は中国地方などの山地(岡山県の恩原高原など)に広がる牧草地にも、人為的に持ち込まれたシラカンバから種子が散布されて、自生状態になっているのが観察されており、奈良県野迫川(のせがわ)村の山林には植栽された小さなシラカンバ林が存在する。
生態
シラカンバは光要求量の多い極陽樹で成長が早く、生態学的には森林生態系の先駆樹種である。一般的に寿命はあまり長くなく、70~80年程度と言われているが、200年ほど生きた個体もあるとされている。
山火事、暴風、雪崩、土砂崩壊、人為的伐採、焼き畑農業等により前植生が破壊されると、風散布により運ばれてきた種子が速やかに発芽する。土壌中の埋土種子も2~3年以内であれば発芽するようである。種子が休眠状態を打破して発芽するには、山火事の熱、あるいは直射日光の熱や赤色光に短時間でも曝される(これを光発芽という)ことが必要である。そのようにして発芽した幼苗は急速に成長し、十数年でシラカンバ林を形成する。北海道には平地にも多くのシラカンバ林が存在するが、それらを観察すると、ほとんどすべてが二次的に成立した森林すなわち二次林である。ただし、森林が破壊されても、下層にチシマザサ、クマイザサ、ミヤコザサ等のササ類が密生している場合は、シラカンバの種子は発芽できず、たとえ発芽してもすぐに枯れてしまう。
近年、北海道では畜産業を廃業する農家が多く、そのため多くの牧草地がソバ畑に変わっているが、そのまま放置された場所も少なくない。そのような放棄牧草地ではオーチャードグラスやティモシー等の西洋牧草の花茎が伸びて開花結実して衰退し、ギシギシ等の広葉雑草が繁茂するが、それらに混じってシラカンバやヤナギ類、オニグルミ、ヤマハンノキなどの種子が飛んできて発芽し、稚苗が生育している。それをさらに放置し続けて十数年も経つと、ヤナギ類、シラカンバ、オニグルミ、ヤマハンノキ等の早成樹種による落葉広葉樹混交林となるが、条件次第ではシラカンバあるいはオニグルミの純林となることもある。因みに、これらの西洋牧草は常に刈り取られたり牛馬に摂食されたりすることにより地際の成長点による栄養成長が続き長く緑色を保っているが、刈り取りや摂食がなくなると生殖成長に移行し、花茎が伸びて結実後に枯れてしまう。湿地帯が排水路整備や地形改変、地表搔き起しで乾燥化した場合も、埋土種子が発芽してシラカンバ林を形成することがある。さらに、カラマツ人工林などでエゾヤチネズミの食害によりカラマツ苗の成長が抑制されると、シラカンバやウダイカンバの実生苗のほうが成長が早いので、カラマツ林にならずにカンバ林になってしまう例も報告されている。北海道の鉄道防雪林では、時折台風によってトドマツの風倒木が発生するが、風倒木を整理した後に飛来したシラカンバの実生苗を刈らずに育てたり、苗木を植えたりして早期の成林を図ることが多い。
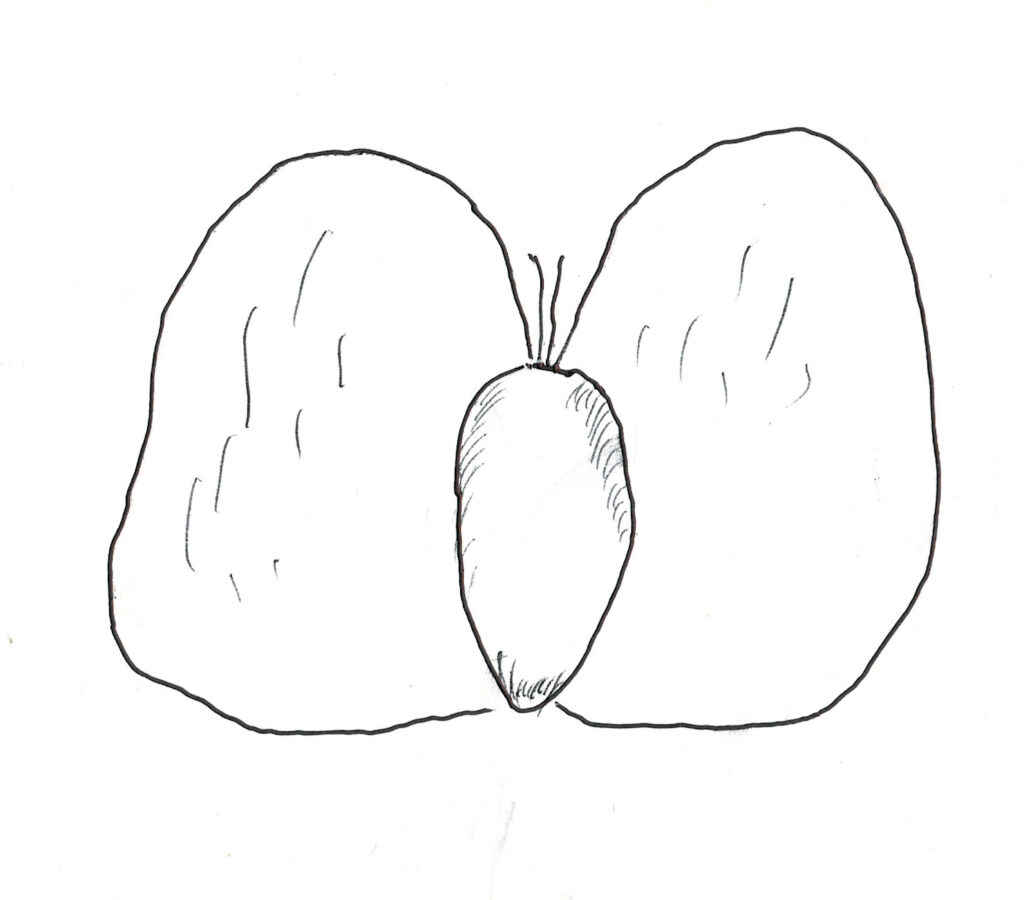
シラカンバは雌雄同株であるが、雄花と雌花は別である。10年生くらいから種子を着け始めるが、その種子(図1)は果穂の脇に沢山結実する。極めて小さく、2枚の翼を持つ翼果であるので風で遠くに飛ばされやすいが、種子には胚乳がなく貯蔵エネルギーはごく僅かしかないので、発芽後はすぐに子葉及び本葉による光合成を盛んに行わなければ成長できない。根は外生菌根を形成し、痩せた土壌でも生育することができる。森林内に落下した種子は赤色光に曝されず、直射日光の熱も得られないので発芽しないが、たとえ発芽したとしても、普通は日照不足ですぐに枯れてしまう。しかし、落下地点が大枝の折損や倒木により林冠に大きなギャップが形成されている場合は、発芽と成長に必要な光が得られ、大きく成長することがある。
結実したシラカンバの種子は、落下した土地の条件によって、その秋のうちに発芽する個体と、翌春に発芽する個体を分ける。秋に発芽する個体は厳冬期に枯死してしまうものが多いが、枯死率は冬の気候条件によって大きく変わる。生き残った実生苗は翌春に発芽するものよりも早く大きく成長する。春になってからの発芽は発芽率が高く、安定的な成林を保証する。
ひとたびシラカンバ林が成立すると、シラカンバ林の林床に落下したシラカンバの種子は成長できず、より耐陰性の高いイタヤカエデ、カエデ類、ミズナラ、コナラ、トドマツ等が次々と侵入してくるので、シラカンバ林と言える期間は短い。
シラカンバの性質
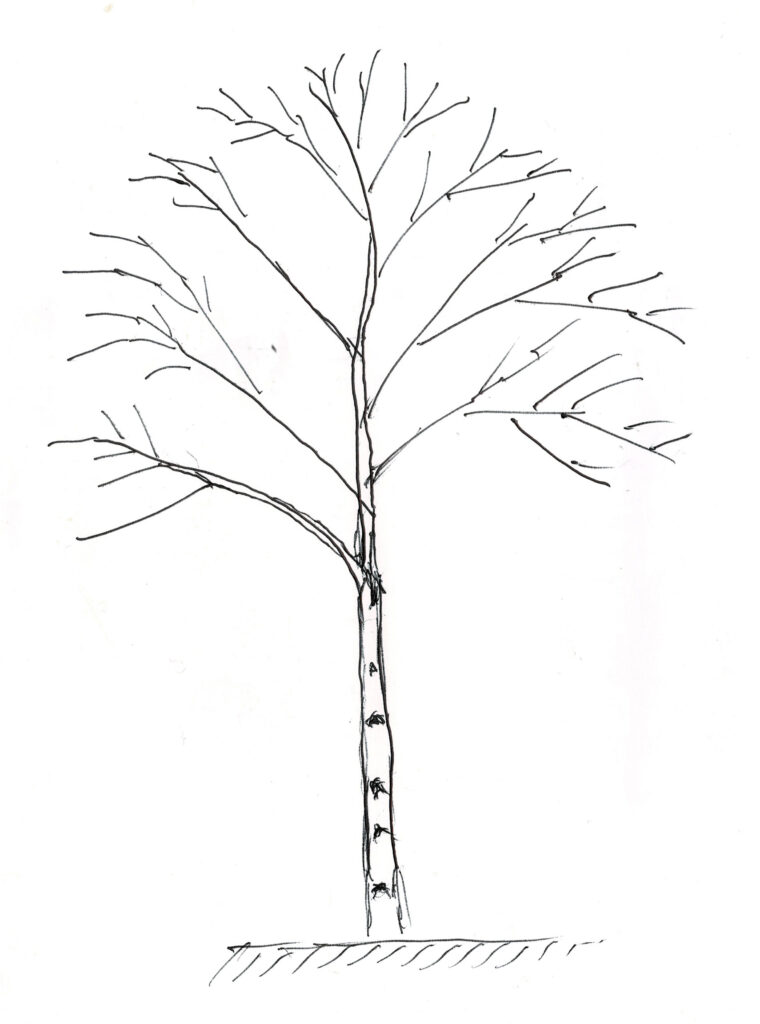
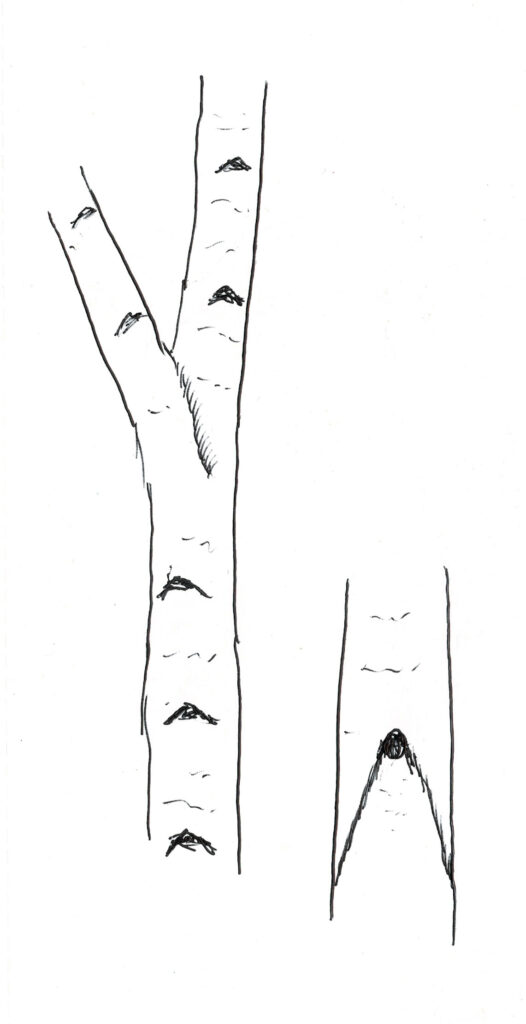
シラカンバは主軸が明瞭で図2のような樹形となるが、幹が少し屈曲しながら成長する。森林では樹冠が小さく、枝下高が極めて高い状態となるが、四方八方、低い位置からも光の来る野原に生育する孤立木の場合、下枝は枯れずに大きな樹冠を形成する。しかし、地面に接する程に枝が低く垂れ下がることはほとんどなく、多くは下枝が徐々に枯れて脱落していく。枝の脱落痕を正面から見ると図3のように白い木肌に黒いブランチバークリッジの痕が極めて明瞭に浮かび上がっている。その様子を英語ではChinese Moustacheと呼んでいる。なお、ダケカンバではこの口髭状のブランチバークリッジの痕跡は不明瞭なので、シラカンバとダケカンバを見分ける特徴の一つとなっている。
シラカンバの樹皮が白いのはベチュリン(Betulin)というトリテルペンの無色透明の粉がコルク層を覆っているからで、小枝にはこのベチュリンがほとんどないので、樹皮は黒い。なお、ベチュリンは近年、その制癌剤としての薬理効果が注目されている。
シラカンバの樹皮は横方向に薄く剥がれる性質があるが、これは横方向に細胞が並んだコルク層が多層構造を示しているからである。樹木の形成層は内側に木部、外側に篩部を形成する。その段階では、木部も篩部も軸方向に細胞が並んでいる。篩部では内側から新しい篩部細胞が形成されると、多くの古い篩部細胞は押し潰されて死滅するが、一部の篩部細胞はコルク形成層に変わり、周皮を形成する。周皮は樹木の肥大成長と共に横方向に引っ張られながらコルク層やコルク皮層の細胞を増していくので、徐々に横方向に引き伸ばされる。それにつれて皮目コルク形成層も横方向に分裂を重ね、皮目が横1列に並ぶ状態になる。そして、ある段階に達するとちぎれて横方向に薄紙のようにコルクが剥がれてくる。樹皮の様子が独特なので、飾り物などの工芸品として利用されている。
樹皮には外皮に蝋物質が分泌され、若い枝には油脂細胞が存在する。そのため燃えやすく、薪ストーブや焚火の着火剤としてよく利用されている。俗説だと思うが、結婚式を意味する「華燭之典」の華燭とは樺(かんば)の樹皮を燃やす松明(たいまつ)のことを意味するとの説もある。また、樹皮を丸めて縛ると水に浮くので、現在も樺太では漁網の「浮き」として使われているようである。カンバ類の樹皮の蝋物質は雨水の浸透を阻止して厳冬期に樹皮細胞が凍結するのを阻止する働きがあると考えられるが、小枝に含まれる油脂成分も、エネルギーを貯蔵する機能と、厳冬期の細胞凍結を防ぐ機能を持つと考えられる。
因みに、カンバの語源は万葉集に出てくる「かには」であろうと言われているが、この「かには」はカンバではなくサクラの樹皮であるという説もある。また、アイヌ語の「かりんぱ」がカンバに変わったという説も出されている。
シラカンバは散孔材で材質は乾くと硬いが軽く腐朽しやすいので、構造材や野外で利用する道具材には不向きである。材は乾燥すると硬くなり、木肌も辺材は淡黄白色、心材は淡灰褐色であるが、境界が不明瞭なことが多い。年輪界が細かく明るい材色なので、その美しさを強調しながら内装材、家具材や調度品に利用されている。心材がやや暗色で辺材との区別が明瞭な場合は、過去に大きく傷ついた時に形成された偽心材の可能性がある。偽心材には正常な心材よりもポリフェノールが多く含まれる傾向がある。
大きな傷害が生じると、その傷口から腐朽が進行しやすいので、強剪定を避け、枯れ枝のみを切除するのが良い。また、樹勢が衰退するとゴマダラカミキリ、ウスバカミキリ、カバノモグリバエ等の穿孔虫の被害を受けやすい。とくにゴマダラカミキリは根元付近に穿孔して大穴を開けるので、根元での腐朽と幹折れの原因となり易い。また、カバノモグリバエやミノドヒラタモグリガの幼虫はピスフレック(材の断面に生じる小さな黒い斑点)の原因となる。ピスフレックは幹の高さ1m以下で発生しやすいとされている。
シラカンバの導管液はイタヤカエデなどから採取されるメープルシロップと同じく早春の一時期に採取される。厳冬期が終わり、葉が展開する前の2週間から3週間ほどの間は、耐凍性を高めるために細胞内に溜め込んでいた糖分を導管のほうに流出させ、また葉が展開する前なので、蒸散力はまだ働かず、根は根圧で水分を吸収しており、幹に穴をあけると、導管液が押し上げるように上昇してくるので導管液が採取できる。この導管液にはほのかな甘みがあり、煮詰めてシロップとして利用されるが、虫歯を予防する効果のあるキシリトール等を含んでおり、肌にも良いので化粧水としても利用されている。
カンバ類の花粉症は日本ではスギ・ヒノキの花粉症の影に隠れてあまり話題にはなっていないが、ロシアや北欧では患者数が極めて多く、北海道でも問題となっている。
参考文献
・岩泉正和(2007)シラカンバ、ダケカンバ、ウダイカンバの林木遺伝資源保存林、林木遺伝資源情報第12-1
・石澤宣夫ほか(2013)シラカンバ人工林におけるピスフレックの発生実態-美唄市の44年生林分での事例-、北方林業61
・大住克博(2003)シラカンバの分布限界地について考える、森林科学39巻
・小山浩正(2002)シラカンバの発芽フェノロジーと適応戦略としての意義、北海道林業試験場研究報告第39号
\ 最新情報をチェック /

